
For volunteers taking part in sleep deprivation studies:
• What is fMRI?
• Is MRI dangerous?
• Are MRI and CT brain scans the same?
• Are there any requirements before I can be scanned in the MRI scanner?
• Is there any preparation I must do before being scanned?
• What is the in-scanner experimental procedure like?
• What am I expected to do in the scanner?
• Are there any prerequisites for taking part in the sleep deprivation studies?
• How long will I be sleep-deprived? What do I do during this time?
• How much will I get paid for my participation?
• I'm interested! How do I take part?
For volunteers taking part in aging or S-LABS studies:
• What is S-LABS?
• Where is S-LABS conducted?
• Am I eligible to take part?
• How long does the procedure take?
• Do I need to prepare anything prior to my appointment date?
• So how much blood are you going to collect?
• Do I need to pay to participate in this study?
• What will I get out of this study?
• Will I be informed of the results of the study?
• Who can I contact to get more information?
For volunteers taking part in sleep deprivation studies:
What is fMRI
fMRI stands for functional magnetic resonance imaging. When we perform a particular cognitive task, neural activity in specific parts of our brains increases. This results in greater need for glucose and oxygen, both of which are met by increasing blood flow. There is a small difference in magnetic susceptibility (a property of molecules) between oxygenated hemoglobin (the oxygen carrying substance in blood) and de-oxygenated hemoglobin. With increased blood flow to activated brain areas, there is a shift in the proportion of oxygenated (increased) to de-oxygenated hemoglobin. This difference can be detected in the scanner; however it is very small. Only by acquiring many samples (images) can we use this difference to obtain statistically-significant brain activation maps.
Is MRI dangerous?
No! fMRI is a non-invasive method of medical imaging that does not involve ionizing radiation like X-rays or CT scans. (See below)
Are MRI and CT brain scans the same?
The recent article in The Straits Times, âDangerous brain scans foundâ should prove little cause for alarm among Magnetic Resonance Imaging (MRI) participants. The dangerous brain scans in question refer to a series of Computed Tomography, or CT brain perfusion scans, administered at Cedar-Sinai Medical Center in Los Angeles and other hospitals. These scans delivered eight times the normal radiation levels of a normal CT perfusion scan, which caused patients to suffer side effects such as hair loss and skin redness. As a result, the Food and Drug Administration is set to impose limitations on future CT scans. | | 
Read the original article |
| These potential dangers that accompany CT scans do not, however, apply to MRI scans even though from the outside, the scanners look the same. The physical phenomena used by CT and MR are very different. Computed Tomography is a medical imaging tool that primarily uses x-ray radiation to create an image. The machine rotates around the body while delivering x-ray beams at multiple points; detectors then measure the radiation absorbed by various tissues and bones. This data is used to generate two dimensional cross-sectional slices of the body. | | 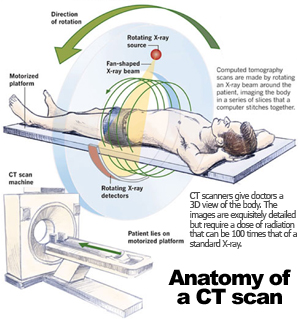
Photo credit: http://www.worldculturepictorial.com |
| In contrast, MRI technology does not rely on ionizing radiation. Rather, an MRI scan uses a constant magnetic field, changing local magnetic fields, and radiofrequency signals to produce images. As long as the proper safety precautions are observed, MRI is an extremely safe procedure. To date, over 150 million patients have undergone MRI, with approximately 10 million patients taking an MRI examination each year. MRI is noninvasive, pain free and causes no known short or long term damage to tissues. | | 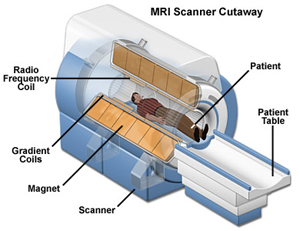
Photo credit: http://www.themesotheliomalibrary.com |
The safety of volunteers is very important to us in our lab. That is why we take care to screen volunteers for possible pregnancy and for metal implants, before allowing them to participate in experiments. These precautions are closely followed by all our scanner personnel, who are well-trained and experienced in detecting any contraindications to MRI scanning. If you have any further questions about MRI or questions about our lab, please feel free to contact us at [email protected].
Are there any requirements before I can be scanned in the MRI scanner?
Participants should be healthy, of average build and should not be claustrophobic. You should not have any metallic implants such as a pacemaker, aneurysm clips or heart valve in your body. You should also not be wearing extensive braces.
Is there any preparation I must do before being scanned?
You should avoid drinking alcohol and caffeinated drinks 24 hours prior to the scan, and abstain from smoking six hours before the scan. We request that participants wear comfortable and loose clothing (such as a T-shirt and bermudas) without embellishments when they come for an experimental session.
What is the in-scanner experimental procedure like?
The in-scanner procedure varies depending on the specific study. Typically there are three parts to the scan. Before and after the experimental task, you can relax while we take structural scans of your brain. The actual experiment may consist of 4-10 runs, taking up to 60 minutes. You will spend about 65 to 90 minutes in total in the scanner at a go.
What am I expected to do in the scanner?
You are required to lie still during the scan, and not move as much as you can. Focus your attention on performing the given task(s) as accurately and as quickly as possible.
Are there any prerequisites for taking part in the sleep deprivation studies?
You should be right-handed, and have no chronic illness or history of sleep disorders. You should also have regular sleeping habits, getting 6.5 to 9 hours of sleep every night, and sleeping before 12:30AM and waking up before 9:00AM daily.
How long will I be sleep-deprived? What do I do during this time?
The time that you will be sleep deprived for varies from study to study, but it is typically a period of 24 hours. You will be asked to come into our lab in the evening, and we will monitor you throughout the night to make sure that you do not fall asleep. During this time, there may be tests and questionnaires that you will be asked to fill in. When you are not being tested, you will be free to read, do your own work, or watch DVDs.
How much will I get paid for my participation?
Payment varies from study to study -- but you can expect to receive between $50 for one session (i.e. one overnight or one morning session) and $120 for completing one overnight and one morning session.
I'm interested! How do I take part?
Please fill up our sleep questionnaire here. If we find you suitable, we will contact you when we begin the recruitment phrase of our sleep deprivation studies.
For volunteers taking part in aging or S-LABS studies:
What is S-LABS?
S-LABS stands for the Singapore-Longitudinal Aging Brain Study. It is a five-year study which will involve about 350 men and women above the age of 55 years. This study will examine the factors that increase your likelihood of thinking clearly and having an active mind well into your advanced years.
Volunteers will undergo the following assessments:
• Magnetic Resonance Imaging (MRI) brain scans
• Cognitive assessments, namely memory tests
• Blood tests
Volunteers will be evaluated three times: in the Year 2005/6, 2007/8 and 2009/10 at 18-month intervals to observe how brain measurements, cognition and health variables change with time.
The first pass of S-LABS was completed in August 2006. We are about to embark on pass 2.
Where is S-LABS conducted?
S-LABS is conducted in the Cognitive Neuroscience Laboratory (CNL), which is located near the Singapore General Hospital. Please refer to our location page for instructions on how to find us.
Am I eligible to take part?
If you are 55 years old and in good health, you are eligible to take part in this study.
How long does each visit take?
You will usually be in our lab for about half a day.
Do I need to prepare anything prior to my appointment date?
For some blood tests, volunteers have to fast on the night before their appointment day. You should consume nothing but plain water from midnight until the blood test is done the following morning.
So how much blood are you going to collect?
We will collect a total of 10-12 ml of blood.
Do I need to pay to participate in this study?
No.
What will I get out of this study?
Your data will provide us with valuable information about aging to future generations. You will be paid $30 for each of the 3 sessions in which you participate.
Will I be informed of the results of this study?
A copy of your blood results will be mailed to you. There will not be any written report on the results of your brain scan; however, you will be informed if abnormalities are found in your brain images.
Who can I contact for more information?
For more information and enquiries, you may call 6326-6175 to speak to any of our friendly S-LABS staff. Thanks!
